

 Membrane proteins account for 25–30% of the human proteome and represent over half of all FDA-approved drug targets1,2. Yet, while producing them for drug discovery, these proteins remain famously fragile during purification. While detergents can extract receptors and transporters, they strip away annular lipids and often push proteins into non-native conformations3.
Membrane proteins account for 25–30% of the human proteome and represent over half of all FDA-approved drug targets1,2. Yet, while producing them for drug discovery, these proteins remain famously fragile during purification. While detergents can extract receptors and transporters, they strip away annular lipids and often push proteins into non-native conformations3.
Nanodiscs solve this problem by keeping a small, soluble patch of lipid bilayer around the protein—turning membrane-protein biochemistry into something closer to “solution biology”.
This guide explains what nanodiscs are, why they outperform detergent micelles, and the practical knobs you can tune (membrane scaffold protein, lipid composition, stoichiometry). We also cover a key shift for fast iteration: using pre-assembled nanodiscs directly in cell-free protein synthesis (CFPS) to enable co-translational insertion.
What are nanodiscs?

Figure 1 | Schematic of a membrane protein stabilized in a nanodisc. The membrane scaffold proteins (MSPs, dark blue) wrap around the phospholipids (light blue), holding the membrane protein (purple) in a native-like bilayer.
Nanodiscs are small (7–16+ nm diameter), discoidal lipid bilayers wrapped by an amphipathic “belt” that shields the hydrophobic edge of the membrane4. The most widely used format is the Membrane Scaffold Protein (MSP) nanodisc, derived from human apolipoprotein A-1.
Two copies of the MSP encircle a patch of phospholipids, generating a synthetic model membrane system. This allows membrane proteins to remain soluble while retaining a native-like lipid environment.
Key Components:
- Phospholipids: Artificial lipids of your choice.
- The Belt (MSP): A protein scaffold that determines the disc size and holds the lipids together.
Nanodisc assembly: Two ways to reconstitute
There are two primary methods to get a membrane protein into a nanodisc: the traditional cell-based “detergent removal” method and the newer cell-free “co-translational” method.
1. The traditional route: Detergent removal
Classically, nanodisc assembly is a multi-step process5. In cell-based membrane protein expression, the proteins are embedded in the host membrane. To purify them, protein scientists typically perform detergent extraction (solubilization into micelles). For nanodisc reconstitution, the detergent-protein solution is mixed with purified MSPs and lipids at a specific stoichiometry and the detergent is slowly removed (typically using hydrophobic beads), forcing the lipids and MSPs to self-assemble around the protein.
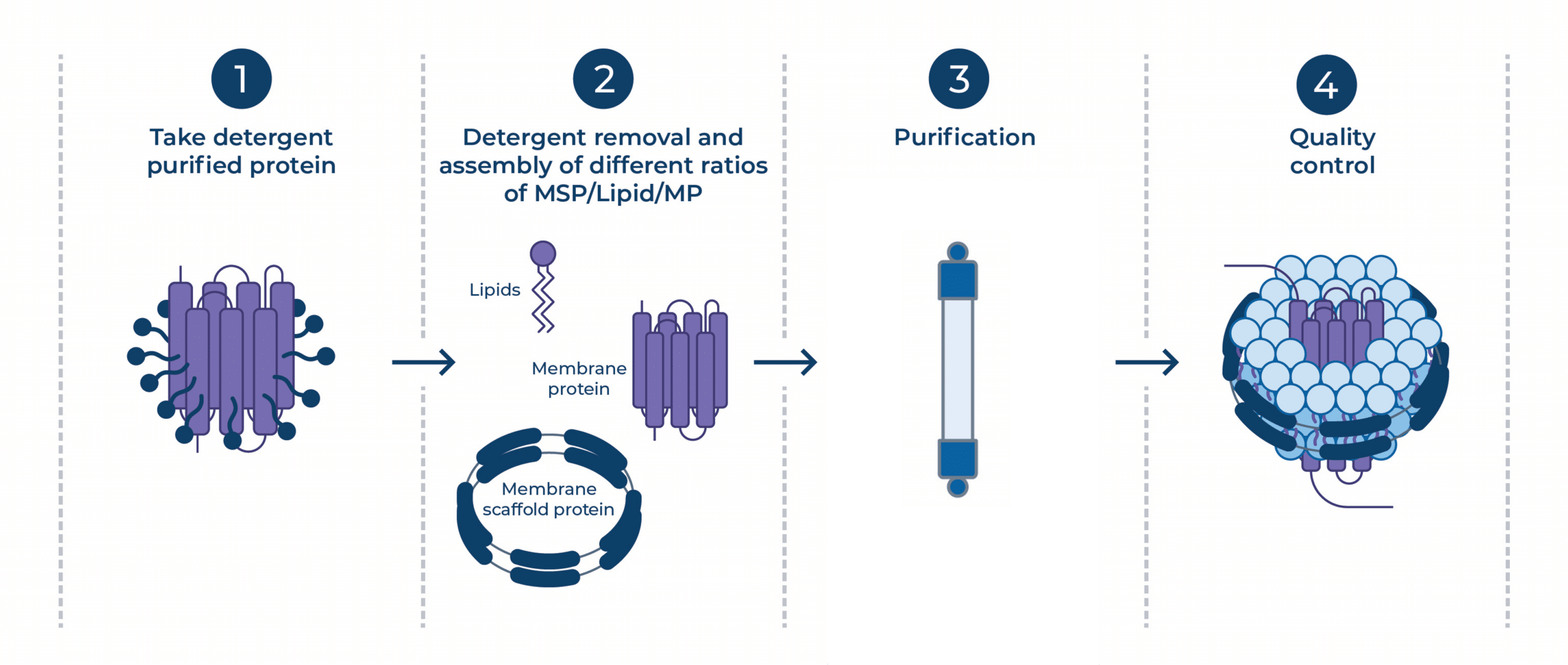
Figure 2 | Schematic overview of nanodisc reconstitution of detergent-solubilized membrane proteins.
The bottleneck: This relies on the protein surviving in a detergent micelle for hours or days. Many fragile targets denature or aggregate during this “detergent window” before they reach the safety of the nanodisc6.
2. The fast track: Co-translational insertion during CFPS
To bypass the instability of detergents, modern workflows increasingly utilize CFPS. By adding pre-assembled “empty” nanodiscs directly into the translation reaction, the nascent protein can insert into the lipid bilayer as it is being synthesized via a translocon-unassisted mechanism (co-translational insertion)7.
This process mimics the natural, ribosome-associated, folding of membrane proteins and allows for rapid screening of different MSP-lipid environments.
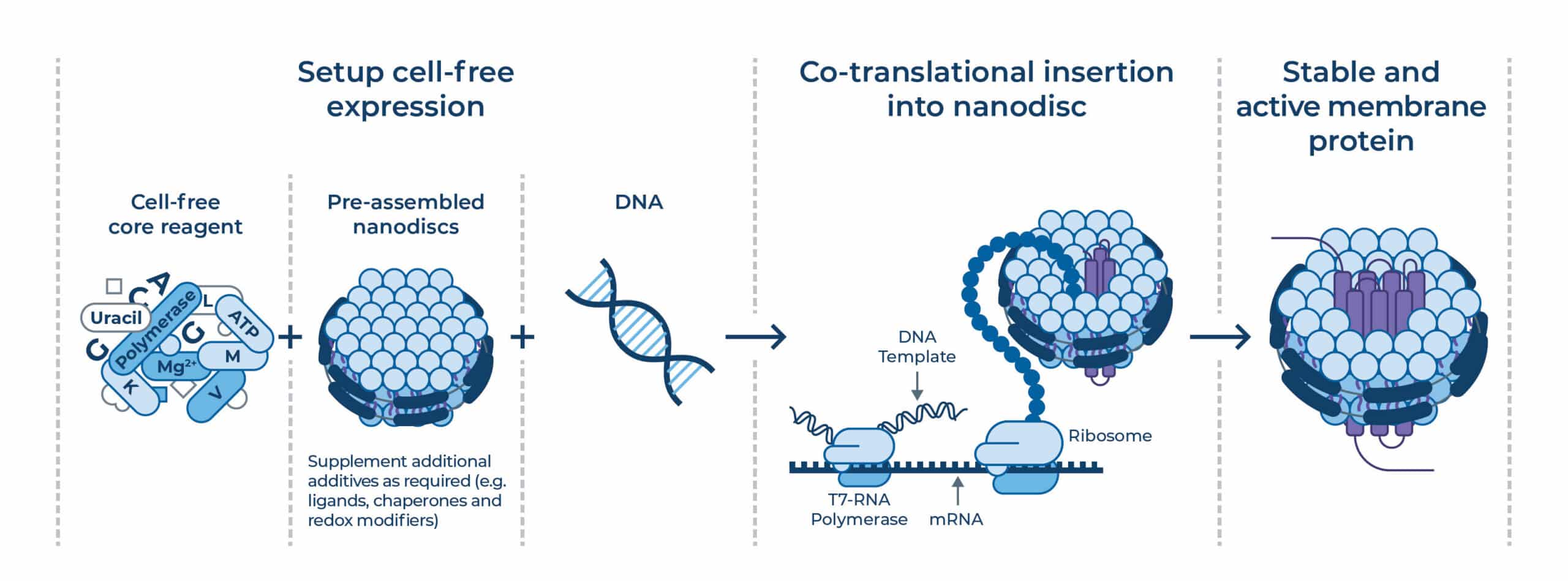
Figure 3 | Combining CFPS and pre-assembled nanodiscs enables co-translational insertion of membrane proteins and faster iteration.
Tunable membranes: What you can optimize in nanodiscs
One of the greatest advantages of nanodiscs is that they are a “tunable” design space. You aren’t stuck with a single condition.
1. Disc size (MSP selection)
The diameter of the nanodisc is controlled by the length of the MSP belt. This is critical because the disc must be large enough to prevent steric crowding that could interfere with protein insertion, folding or multimeric assembly. However, the diameter must also be small enough to maintain homogeneity to prevent the incorrect number of proteins from inserting into one nanodisc.
Evidence: Studies on Endothelin A and B receptors (ETAR and ETBR) showed that expressing them in MSP1E3D1 nanodiscs (~13 nm) produced significantly higher yields of stable protein compared to the MSP1D1 (~9.5 nm) discs9.
Table 1 | Typical MSP nanodisc sizes.
| MSP example | Diameter | Use case |
|---|---|---|
| MSP1D1ΔH5 | ~8 nm | Many single-pass proteins. |
| MSP1D1 | ~9–10 nm | Single-pass proteins; G-protein coupled receptors (GPCRs). |
| MSP1E3D1 | ~12–13 nm | GPCRs; multi-pass proteins; larger complexes. |
| MSP2N2 | ~16–17 nm | Large oligomeric complexes. |
2. Lipid composition
You aren’t limited to a single “standard” lipid. Changing the chemistry alters the physical forces on your protein:
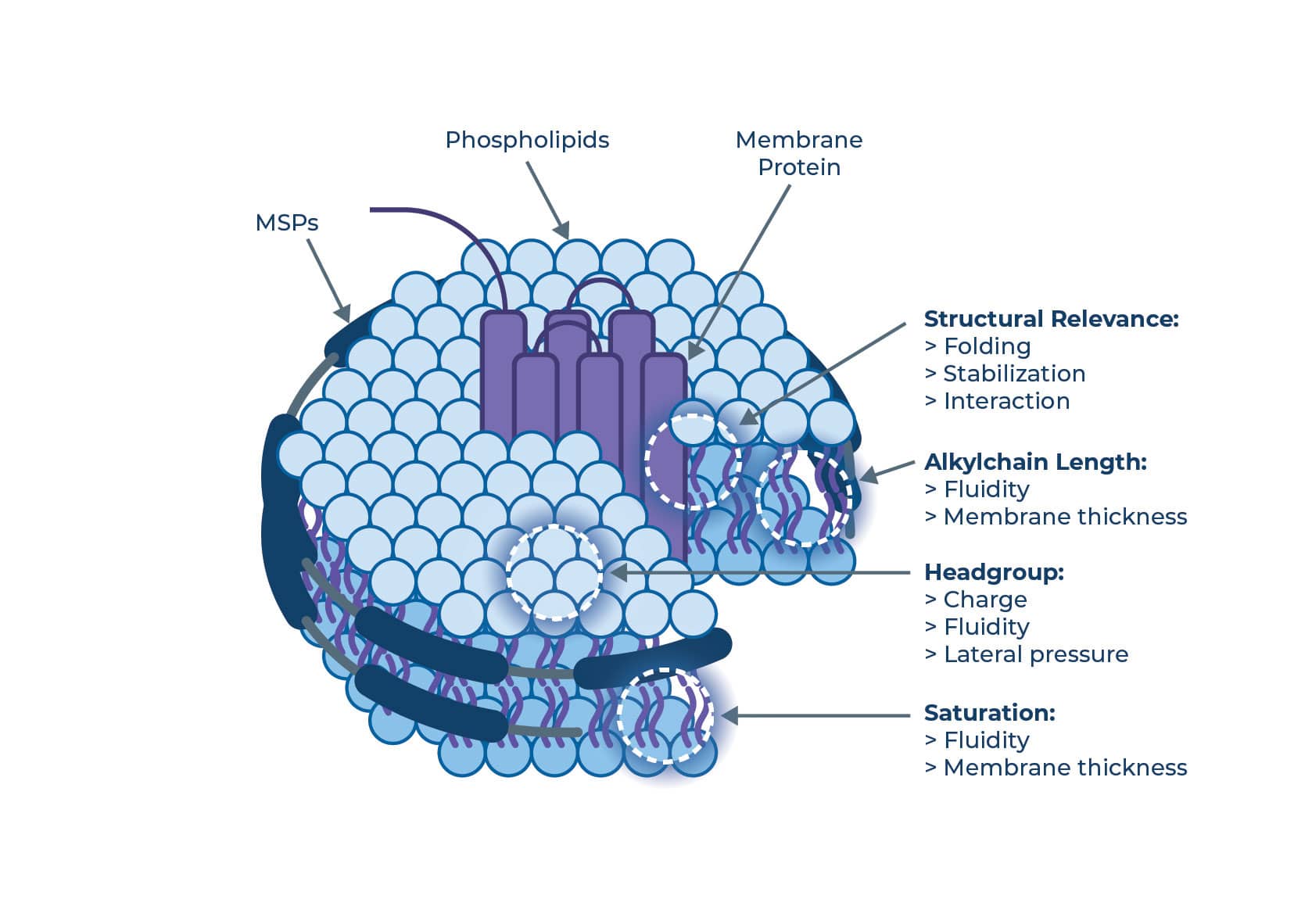
Figure 4 | Lipid architecture influences physical forces such as membrane thickness, fluidity, and charge interactions.
Alkyl chain length: Varying the carbon chain length (e.g., C14 vs. C18) changes the membrane thickness.
Saturation: The number of double bonds determines chain flexibility and membrane fluidity. Saturated lipids form tighter, more ordered packing. A more fluid membrane can enable conformational changes, while a rigid one might stabilize a specific state.
Headgroup charge: The headgroup charge is crucial for mimicking native membranes, and contributes to the stability and functional conformation of many membrane proteins.
Evidence:
-
- Charge matters: The β1-adrenergic receptor (β1AR) showed a 12-fold increase in activity when reconstituted in negatively charged lipids (like PG (phosphatidylglycerol)) compared to neutral lipids (PC (phosphatidylcholine))9.
- Saturation:β1AR and ETBR activity increased using fatty acids of longer 18 carbon chain lengths, compared to 14 carbon, and with a flexible trans configuration8–11.
- Cardiolipin: Including 20% cardiolipin in DOPG (Dioleoylphosphatidylglycerol) nanodiscs increased the active fraction of human β1AR by 20%11.
- Cholesterol & complex mixes: Functional screens suggest that “physiological” mixtures (e.g., 20% POPS (1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-L-serine), 8% cholesterol, 72% POPC (1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine)) can provide better GPCR enrichment than simple PC formulations12. Cholesterol specifically can influence signaling by binding directly to the protein or altering membrane fluidity.
Quality control: How to know your nanodiscs are working
Regardless of whether you use the cell-based detergent removal approach or CFPS, you need to verify your membrane-protein embedded nanodiscs.
- Did it incorporate?
- Is it monodisperse?
- Is it functional?
SEC (Size Exclusion Chromatography) is the standard “first gate”. SEC can be used to identify and separate out empty nanodiscs and aggregates from the protein-loaded nanodiscs.
Evidence: SEC of CFPS-made β1AR, revealed two fractions11. The lower molecular weight fraction contained monomeric receptor-nanodisc complexes with 40–60% active protein, whereas the high-molecular-weight aggregate fraction had <5% activity.

Figure 5 | SEC provides a rapid QC gate for monodispersity and enables purification of “active” fraction before committing to functional assays. 1 represents a non-active, aggregated protein. 2 corresponds to a correctly stabilized, active protein embedded within the nanodisc
Applications: Where nanodiscs add the most value
Nanodiscs have become essential across membrane protein science because they bridge the gap between native physiology and structural accessibility.
1. Cryo-EM: Solving the “preferred orientation” problemMembrane proteins in detergents often exhibit preferred orientations (e.g., top-down view) in the vitreous ice, making it difficult to collect enough side-view images for a 3D reconstruction. Nanodiscs have a distinct, rigid disc shape that tends to orient more randomly, providing better angular coverage in the ice13. This often improves particle distribution and allows researchers to resolve high-resolution structures. Nanodiscs can also reduce aggregation, preserve target function and structure, and provide immunity to freezing and mounting.
Example: Human histamine 2 receptor (H2R) in an active conformation with bound histamine and in complex with Gs heterotrimeric protein at an overall resolution of 3.4 Å using MSP1E3D1-DOPG19. Transient receptor potential vanilloid 1 (TRPV1) ion channel in the un-liganded, agonist-bound and antagonist-bound states at resolutions of 3.2, 2.9 and 3.4 Å, respectively14.
2. Nuclear magnetic resonance (NMR) spectroscopyNanodiscs (especially ~8–10 nm discs) are small enough to tumble effectively in solution, enabling high-resolution structural studies in a bilayer15.Example: A high-resolution NMR structure of bacteriorhodopsin and Outer Membrane Protein X (OmpX)16 using small nanodiscs, highly deuterated proteins and lipids, and advanced sampling methods.
3. GPCR pharmacology & drug discoveryNanodiscs enable detergent-free binding assays (SPR, DEL, Radioligand) where the receptor is stable. They are particularly useful for fragment screening, which requires long incubation times that detergent-solubilized proteins cannot withstand.
Nanodiscs vs. detergents (quick decision guide)
- Choose detergents if your protein is robust and you need a quick, cheap extraction.
- Choose MSP nanodiscs if you need high stability, functional assays, or structural data (Cryo-EM), or are using CFPS.
| Feature | Detergent Micelles | MSP Nanodiscs |
|---|---|---|
 | | |
| Structure | Surfactant micelle (non-bilayer) | True planar lipid bilayer |
| Stability | Low (degrades minutes to hours)6,11,17 | High (Stabilized by belt protein and retains activity for days to weeks)6,11,17 |
| Annular Lipids | Stripped away | Reconstituted (User defined) |
| Key Advantage | Low cost; standard for extraction. | Very customizable; protein stability; best for Cryo-EM & CFPS. |
| Key Limitation | Destabilizes fragile targets; non-native environment. | Assembly can be complex (unless using CFPS). |
Limitations of nanodiscs
While powerful, nanodiscs are not a universal solution. It is important to consider these constraints:
- Fixed size constraints: Unlike liposomes, which can expand, a nanodisc has a rigid boundary defined by the MSP belt. If a protein undergoes a large conformational change or oligomerization that exceeds the disc diameter, it may be destabilized. Note: Selecting an MSP of an appropriate, larger diameter can be used to resolve these issues. Covalently circularized nanodiscs (cND) and DNA corralled nanodiscs (DCND) have opened up the possibility of engineering nanodiscs of size up to 90 nm18.
- Side access blocked: The MSP belt covers the hydrophobic rim of the bilayer. If your drug or ligand enters the protein via the lipid bilayer (lateral diffusion), the belt protein might physically block this access path. Note: Most drug targets bind ligands via accessible extracellular or intracellular domains, making this a rare concern for standard pharmacology.
- Assembly complexity: Traditional assembly requires precise optimization of lipid-to-protein ratios. If the ratio is off, you may end up with empty discs or aggregates. Note: This complexity is largely mitigated by using pre-assembled discs in CFPS workflows.
Accelerating discovery with Nuclera
MSP-lipid nanodiscs offer flexibility, with many ways to optimize them for your target membrane protein. Membrane proteins rarely fail because a single variable is “wrong”—they fail because exploring the matrix of constructs, lipids, and additives is too slow. Combining nanodiscs with CFPS, you can rapidly identify the best conditions for your protein.
Nuclera’s eProtein Discovery™ system allows you to multiplex screen panels of constructs, nanodiscs, and additional additives (redox, chaperones, etc.) in a cell-free environment using automated digital microfluidics. Start with a 24-hour multiplex screen, and decide which are the best conditions to scale up for assay-ready membrane proteins in days rather than months.
Paired with Nuclera’s validated GPCR-specific nanodiscs, ready to plug directly into your eProtein Discovery™ system, you can eliminate the variability of homemade formulations and screen the specific charge, fluidity, and cholesterol needs for GPCR activity and yield. Our panel covers the critical design space necessary to deliver purified, assay-ready GPCRs in just 48 hours.
References
- Krogh, A. et al. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580 (2001).
- Overington, J. P., Al-Lazikani, B. & Hopkins, A. L. How many drug targets are there? Nat. Rev. Drug Discov. 5, 993–996 (2006).
- Seddon, A. M., Curnow, P. & Booth, P. J. Membrane proteins, lipids and detergents: not just a soap opera. Biochim. Biophys. Acta 1666, 105–117 (2004).
- Denisov, I. G., Grinkova, Y. V., Lazarides, A. A. & Sligar, S. G. Directed self-assembly of monodisperse phospholipid bilayer Nanodiscs with controlled size. J. Am. Chem. Soc. 126, 3477–3487 (2004).
- Ritchie, T. K. et al. Chapter 11 – Reconstitution of membrane proteins in phospholipid bilayer nanodiscs. Methods Enzymol. 464, 211–231 (2009).
- Bayburt, T. H. & Sligar, S. G. Membrane protein assembly into Nanodiscs. FEBS Lett. 584, 1721–1727 (2010).
- Roos, C. et al. High-level cell-free production of membrane proteins with nanodiscs. Methods Mol. Biol. 1118, 109–130 (2014).
- Rues, R.-B. et al. Systematic optimization of cell-free synthesized human endothelin B receptor folding. Methods 147, 73–83 (2018).
- Rues, R. B., Dötsch, V. & Bernhard, F. Co-translational formation and pharmacological characterisation of β₁-adrenergic receptor–nanodisc complexes with different lipid environments. Biochim. Biophys. Acta 1858, 1306–1316 (2016).
- Proverbio, D., Roos, C., Beyermann, M., Orbán, E., Dötsch, V. & Bernhard, F. Functional properties of cellfree expressed human endothelin A and endothelin B receptors in artificial membrane environments. Biochim. Biophys. Acta (BBA) Biomembranes 1828, 2182–2192 (2013).
- Köck, Z., Ermel, U., Martin, J., Morgner, N., Frangakis, A. S., Dötsch, V., Hilger, D. & Bernhard, F. Biochemical characterisation of cell-free synthesized human β₁-adrenergic receptor cotranslationally inserted into nanodiscs. J. Mol. Biol. 434, 167687 (2022).
- Roy, J., Pondenis, H., Fan, T. M. & Das, A. Direct capture of functional proteins from mammalian plasma membranes into nanodiscs. Biochemistry 54, 6299–6302 (2015).
- Denisov, I. G. & Sligar, S. G. Nanodiscs in membrane biochemistry and biophysics. Chem. Rev. 117, 4669–4713 (2017).
- Gao, Y., Cao, E., Julius, D. & Cheng, Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action. Nature 534, 347–351 (2016).
- Glück, J. M. et al. Nanodiscs allow high-resolution NMR of membrane proteins. J. Am. Chem. Soc. 131, 12060–12061 (2009).
- Hagn, F., Etzkorn, M., Raschle, T. & Wagner, G. Optimized phospholipid bilayer nanodiscs facilitate high-resolution structure determination of membrane proteins. J. Am. Chem. Soc. 135, 1194–1197 (2013).
- Harding, P. J. et al. Constitutive dimerization of the G-protein coupled receptor, neurotensin receptor 1, reconstituted into phospholipid bilayers. Biophys. J. 96, 964–973 (2009).
- Das, K. M. P., Shih, W. M., Wagner, G. & Nasr, M. L. Large nanodiscs: a potential game changer in structural biology of membrane protein complexes and virus entry. Front. Bioeng. Biotechnol. 8, 539 (2020).
- Köck, Z. et al. Cryo-EM structure of cell-free synthesized human histamine 2 receptor/Gs complex in nanodisc environment. Nat. Commun. 15, 1831 (2024).


